Calponin 2
| CNN2 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||
| Identifiers | |||||||||||||||||
| Aliases | CNN2 | ||||||||||||||||
| External IDs | MGI: 105093 HomoloGene: 3215 GeneCards: CNN2 | ||||||||||||||||
| |||||||||||||||||
| Orthologs | |||||||||||||||||
| Species | Human | Mouse | |||||||||||||||
| Entrez | |||||||||||||||||
| Ensembl | |||||||||||||||||
| UniProt | |||||||||||||||||
| RefSeq (mRNA) | |||||||||||||||||
| RefSeq (protein) | |||||||||||||||||
| Location (UCSC) | Chr 19: 1.03 – 1.04 Mb | Chr 10: 79.99 – 80 Mb | |||||||||||||||
| PubMed search | [1] | [2] | |||||||||||||||
| Wikidata | |||||||||||||||||
| View/Edit Human | View/Edit Mouse |
Calponin 2 is a protein that in humans is encoded by the CNN2 gene.
The CNN2 gene is located at 19p13.3 in the human chromosomal genome,[3] encoding the protein calponin 2. Calponin 2 is one of the three isoforms of calponin and an actin filament-associated regulatory protein with wide tissue distributions. Human calponin 2 is a 33.7-kDa protein consisting of 309 amino acids with an isoelectric point (pI) of 7.23. Accordingly, it is also known as neutral calponin.
Evolution
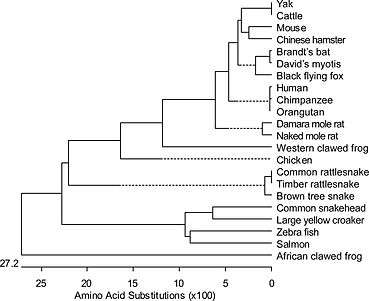
Calponin isoforms are conserved proteins whereas calponin 2 has diverged from calponin 1 and calponin 3 mainly in the C-terminal variable region. Phylogenetic lineage of calponin 2 showed that calponin 2 is conserved among mammalian species but more diverged among amphibian, reptile and fish species (Fig 1).
Tissue distribution
CNN2 is expressed in a broader range of tissue and cell types, including developing and remodeling smooth muscle as well as adult mature smooth muscle,[4] epidermal keratinocytes,[5] fibroblasts,[6] lung alveolar cells,[7] endothelial cells,[8] myeloid white blood cells,[9] platelet,[10] B lymphocyte,[11][12] and myoblasts.[13] These cell types can be classified as a) cells that are physiologically under high mechanical tension (e.g., smooth muscle in the wall of hollow organs, epithelial and endothelial cells), b) cells that have high rates of proliferation (e.g., myoblasts), and c) cells that are actively migrating (e.g., fibroblasts and macrophages). Therefore, the tissue distributions of calponin 2 imply its potential role in regulating cytoskeleton functions and cell motility.[14]
Interaction with other proteins
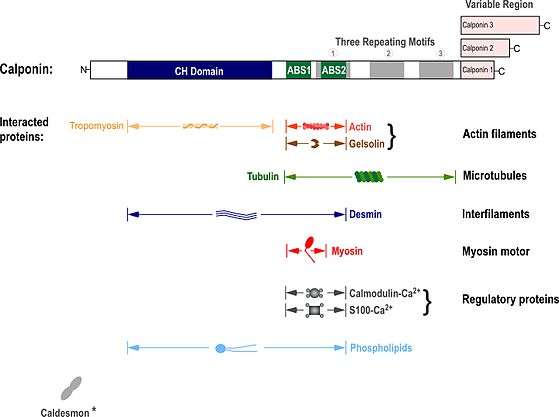
In vitro protein binding studies have demonstrated that calponin binds actin[15] and cross-links actin filaments.[16] Calponin also binds tropomyosin,[17][18] tubulin,[19] desmin,[20][21] Ca2+-calmodulin,[15] Ca2+-S100,[22] myosin,[23] and phospholipids.[24] Calponin also interacts with caldesmon[25] and a-actinin,[26] which however may only reflect their co-localization on actin filaments.[16][27] The variable C-terminal segment regulates actin-binding affinity, and calponin 2 is shown to have the lowest affinity for F-actin among the three isoforms.[28]
Function
Cell proliferation
Significant amounts of calponin 2 are found in growing smooth muscle tissues such as embryonic stomach and urinary bladder as well as the uterus during early pregnancy.[4] The expression of calponin 2 decreases to lower levels in quiescent adult smooth muscle cells while the expression of calponin 1 is up-regulated.[4] Transfective over-expression of calponin 2 inhibited cell proliferation.[4][12] A hypothesis is that higher level of calponin 2 is required in fast proliferating cells to maintain the dynamic equilibrium of the actin cytoskeleton.
Cell motility
Primary fibroblasts and peritoneal macrophages isolated from Cnn2 knockout mice migrate faster than that of wild type control cells.[9] Calponin 2 may affect cell migration differently in different cell types and in different biological processes. A study showed that forced expression of calponin 2 in endothelial cells enhanced angiogenic cell migration in vivo and anti-sense calponin 2 RNA reduced chemotaxis of human umbilical vein endothelial cells in culture.[8] A hypothesis is that a proper level of calponin 2 may be required to maintain the physiological motility of different cell types in different biological processes. Calponin 2’s regulation of cell motility is based on inhibition of actin activated myosin motor function, as fibroblasts isolated from Cnn2 knockout mice showed increased cell traction force generated by myosin II motors.
Cell adhesion
A significant level of calponin 2 is found in human and mouse platelets.[10] Platelet adhesion is a critical step in blood coagulation and thrombosis. In a microfluidic flow-based thrombosis assay, the time to initiation of rapid platelet/thrombus accumulation was significantly longer in blood samples from Cnn2 knockout versus wild type mice.[10] The effect of calponin 2 on facilitating the velocity of cell adhesion was also shown with prostate cancer cells expression high or low levels of calponin 2.[8]
Immune cells
Significant amounts of calponin 2 are found in blood cells of myeloid lineage. Monocytes derived from Cnn2 gene knockout mice proliferated faster than wild type control cells. Calponin 2-null macrophages migrated faster and exhibit enhanced phagocytosis.[9] In global as well as myeloid cell-specific Cnn2 knockout mice, the development of inflammatory arthritis induced by anti-glucose-6-phosphate isomerase serum was significantly attenuated as compared with that in wild type mice . Deletion of calponin 2 in macrophages also significantly attenuated the development of atherosclerosis lesions in apolipoprotein E knockout (ApoE-/-) mice [14]
Regulation by mechanical tension
Gene expression
The expression of calponin 2 is significantly increased in cells cultured on hard versus soft gel substrates that produce high or low traction force and cytoskeleton tension.[6] The expression of calponin 2 in NIH/3T3 cells was decreased when cytoskeleton tension was reduced after blebbstatin inhibition of myosin II motors.[7] To demonstrate the Cnn2 promoter-specific regulation, transfective expression of calponin 2 using a cytomegalovirus promoter was independent of the stiffness of culture substrate.[6]
A binding site for transcriptional factor HES-1 (hairy and enhancer of split 1) has been identified in the 5’-upstream region of mouse Cnn2 promoter, responsible for the mechanical regulation.[13] HES-1 is known to function downstream of the Notch-RBP J signaling pathway,[29] which has been suggested to mediate cellular mechanoregulations.[30][31] Deletion or mutation of the HES-1 site abolished the mechanical regulation and resulted in a substrate stiffness independent high level of transcription. Therefore, the regulatory mechanism is a low tension-induced repression. Corresponding to the down-regulation of Cnn2 gene expression, the level of HES-1 increased in cells cultured on soft gel substrate in comparison with that in cells cultured on hard substrates.[13]
Degradation
Calponin 2 is also regulated by mechanical tension at the protein level. A rapid and selective degradation of calponin 2 occurs in lung tissues after a short period of deflation.[7] This low cytoskeleton tension-induced degradation of calponin 2 in collapsed lung was completely prevented in post mortem mouse lung simply by air inflation to maintain tension applied to the alveolae.[7] The cytoskeleton tension-dependent stability of calponin 2 was further confirmed in monolayer cells cultured on expanded elastic membrane by its rapid degradation after a reduction of the dimension of the cultural substrate to acutely reduce cytoskeleton tension.[7]
References
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ Masuda H, Tanaka K, Takagi M, Ohgami K, Sakamaki T, Shibata N, Takahashi K (1996). "Molecular cloning and characterization of human non-smooth muscle calponin". Journal of Biochemistry. 120 (2): 415–24. doi:10.1093/oxfordjournals.jbchem.a021428. PMID 8889829.
- 1 2 3 4 Hossain MM, Hwang DY, Huang QQ, Sasaki Y, Jin JP (2003). "Developmentally regulated expression of calponin isoforms and the effect of h2-calponin on cell proliferation". American Journal of Physiology. Cell Physiology. 284 (1): C156–67. doi:10.1152/ajpcell.00233.2002. PMID 12388067.
- ↑ Fukui Y, Masuda H, Takagi M, Takahashi K, Kiyokane K (1997). "The presence of h2-calponin in human keratinocyte". Journal of Dermatological Science. 14 (1): 29–36. doi:10.1016/s0923-1811(96)00545-2. PMID 9049805.
- 1 2 3 Hossain MM, Crish JF, Eckert RL, Lin JJ, Jin JP (2005). "h2-Calponin is regulated by mechanical tension and modifies the function of actin cytoskeleton". The Journal of Biological Chemistry. 280 (51): 42442–53. doi:10.1074/jbc.M509952200. PMC 1405912
 . PMID 16236705.
. PMID 16236705. - 1 2 3 4 5 Hossain MM, Smith PG, Wu K, Jin JP (2006). "Cytoskeletal tension regulates both expression and degradation of h2-calponin in lung alveolar cells". Biochemistry. 45 (51): 15670–83. doi:10.1021/bi061718f. PMC 1764619. PMID 17176089.
- 1 2 3 Tang J, Hu G, Hanai J, Yadlapalli G, Lin Y, Zhang B, Galloway J, Bahary N, Sinha S, Thisse B, Thisse C, Jin JP, Zon LI, Sukhatme VP (2006). "A critical role for calponin 2 in vascular development". The Journal of Biological Chemistry. 281 (10): 6664–72. doi:10.1074/jbc.M506991200. PMID 16317011.
- 1 2 3 Huang QQ, Hossain MM, Wu K, Parai K, Pope RM, Jin JP (2008). "Role of H2-calponin in regulating macrophage motility and phagocytosis". The Journal of Biological Chemistry. 283 (38): 25887–99. doi:10.1074/jbc.M801163200. PMC 2533796. PMID 18617524.
- 1 2 3 Hines PC, Gao X, White JC, D'Agostino A, Jin JP (2014). "A novel role of h2-calponin in regulating whole blood thrombosis and platelet adhesion during physiologic flow". Physiological Reports. 2 (12). doi:10.14814/phy2.12228. PMC 4332209. PMID 25472609.
- ↑ Flemming A, Huang QQ, Jin JP, Jumaa H, Herzog S (2015). "A Conditional Knockout Mouse Model Reveals That Calponin-3 Is Dispensable for Early B Cell Development". Plos One. 10 (6): e0128385. doi:10.1371/journal.pone.0128385. PMC 4457629. PMID 26046660.
- 1 2 Moazzem Hossain M, Wang X, Bergan RC, Jin JP (2014). "Diminished expression of h2-calponin in prostate cancer cells promotes cell proliferation, migration and the dependence of cell adhesion on substrate stiffness". FEBS Open Bio. 4: 627–36. doi:10.1016/j.fob.2014.06.003. PMC 4141211. PMID 25161871.
- 1 2 3 Jiang WR, Cady G, Hossain MM, Huang QQ, Wang X, Jin JP (2014). "Mechanoregulation of h2-calponin gene expression and the role of Notch signaling". The Journal of Biological Chemistry. 289 (3): 1617–28. doi:10.1074/jbc.M113.498147. PMC 3894341. PMID 24285540.
- 1 2 Liu R, Jin JP (2015). "Calponin: A mechanical tension-modulated regulator of cytoskeleton and cell motility.". Current Topics in Biochemical Research. 16: 1–15.
- 1 2 Takahashi K, Hiwada K, Kokubu T (1986). "Isolation and characterization of a 34,000-dalton calmodulin- and F-actin-binding protein from chicken gizzard smooth muscle". Biochemical and Biophysical Research Communications. 141 (1): 20–6. doi:10.1016/s0006-291x(86)80328-x. PMID 3606745.
- 1 2 Leinweber B, Tang JX, Stafford WF, Chalovich JM (1999). "Calponin interaction with alpha-actinin-actin: evidence for a structural role for calponin". Biophysical Journal. 77 (6): 3208–17. doi:10.1016/S0006-3495(99)77151-1. PMC 1289132. PMID 10585942.
- ↑ Takahashi K, Abe M, Hiwada K, Kokubu T (1988). "A novel troponin T-like protein (calponin) in vascular smooth muscle: interaction with tropomyosin paracrystals". Journal of Hypertension Supplement. 6 (4): S40–3. doi:10.1097/00004872-198812040-00008. PMID 3241227.
- ↑ Childs TJ, Watson MH, Novy RE, Lin JJ, Mak AS (1992). "Calponin and tropomyosin interactions". Biochimica et Biophysica Acta. 1121 (1-2): 41–6. doi:10.1016/0167-4838(92)90334-a. PMID 1599949.
- ↑ Fujii T, Koizumi Y (1999). "Identification of the binding region of basic calponin on alpha and beta tubulins". Journal of Biochemistry. 125 (5): 869–75. doi:10.1093/oxfordjournals.jbchem.a022362. PMID 10220577.
- ↑ Wang P, Gusev NB (1996). "Interaction of smooth muscle calponin and desmin". FEBS Letters. 392 (3): 255–8. doi:10.1016/0014-5793(96)00824-1. PMID 8774856.
- ↑ Mabuchi K, Li B, Ip W, Tao T (1997). "Association of calponin with desmin intermediate filaments". The Journal of Biological Chemistry. 272 (36): 22662–6. doi:10.1074/jbc.272.36.22662. PMID 9278424.
- ↑ Fujii T, Oomatsuzawa A, Kuzumaki N, Kondo Y (1994). "Calcium-dependent regulation of smooth muscle calponin by S100". Journal of Biochemistry. 116 (1): 121–7. PMID 7798169.
- ↑ Szymanski PT, Tao T (1997). "Localization of protein regions involved in the interaction between calponin and myosin". The Journal of Biological Chemistry. 272 (17): 11142–6. PMID 9111011.
- ↑ Bogatcheva NV, Gusev NB (1995). "Interaction of smooth muscle calponin with phospholipids". FEBS Letters. 371 (2): 123–6. doi:10.1016/0014-5793(95)00868-a. PMID 7672110.
- ↑ Graceffa P, Adam LP, Morgan KG (1996). "Strong interaction between caldesmon and calponin". The Journal of Biological Chemistry. 271 (48): 30336–9. doi:10.1074/jbc.271.48.30336. PMID 8939993.
- ↑ North AJ, Gimona M, Cross RA, Small JV (1994). "Calponin is localised in both the contractile apparatus and the cytoskeleton of smooth muscle cells". Journal of Cell Science. 107 ( Pt 3): 437–44. PMID 8006064.
- ↑ Czuryło EA, Kulikova N, Dbrowska R (1997). "Does calponin interact with caldesmon?". The Journal of Biological Chemistry. 272 (51): 32067–70. doi:10.1074/jbc.272.51.32067. PMID 9405402.
- ↑ Burgstaller G, Kranewitter WJ, Gimona M (2002). "The molecular basis for the autoregulation of calponin by isoform-specific C-terminal tail sequences". Journal of Cell Science. 115 (Pt 10): 2021–9. PMID 11973344.
- ↑ Kageyama R, Ishibashi M, Takebayashi K, Tomita K (1997). "bHLH transcription factors and mammalian neuronal differentiation". The International Journal of Biochemistry & Cell Biology. 29 (12): 1389–99. doi:10.1016/s1357-2725(97)89968-2. PMID 9570134.
- ↑ Morrow D, Sweeney C, Birney YA, Cummins PM, Walls D, Redmond EM, Cahill PA (2005). "Cyclic strain inhibits Notch receptor signaling in vascular smooth muscle cells in vitro". Circulation Research. 96 (5): 567–75. doi:10.1161/01.RES.0000159182.98874.43. PMID 15705961.
- ↑ Morrow D, Sweeney C, Birney YA, Guha S, Collins N, Cummins PM, Murphy R, Walls D, Redmond EM, Cahill PA (2007). "Biomechanical regulation of hedgehog signaling in vascular smooth muscle cells in vitro and in vivo". American Journal of Physiology. Cell Physiology. 292 (1): C488–96. doi:10.1152/ajpcell.00337.2005. PMID 16943241.