Palaeontinidae
| Palaeontinidae Temporal range: Rhaetian to Early Aptian | |
|---|---|
 | |
| Prolystra lithographica | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hemiptera |
| Suborder: | Auchenorrhyncha |
| Infraorder: | Cicadomorpha |
| Superfamily: | †Palaeontinoidea |
| Family: | †Palaeontinidae Handlirsch, 1906 |
| Type genus | |
| †Palaeontina Butler, 1873 | |
| Genera | |
|
See text | |
| Synonyms | |
|
†CicadomorphidaeEvans, 1956 | |
Palaeontinidae, commonly known as giant cicadas, is an extinct family of cicadomorphs. They existed during the Mesozoic era of Europe, Asia, and South America.[1] The family contains around 30 to 40 genera and around a hundred species.[2]
Discovery
The first palaeontinid discovered was Palaeontina oolitica. It consisted of a single forewing[3] collected from the Taynton Limestone Formation (Stonesfield Slate) of Oxfordshire, England by the English natural historian Edward Charlesworth. It was first described in 1873 by the English entomologist Arthur Gardiner Butler in his book Lepidoptera Exotica; or, Descriptions and Illustrations of Exotic Lepidoptera. Butler claimed that it was the oldest butterfly ever recovered, having mistakenly identified it as a butterfly of the family Nymphalidae.[4]
Description and paleobiology
Palaeontinids had large bodies covered with bristles (setae). They had small heads and broad wings. They superficially resemble moths.[5][6] Large palaeontinids like Colossocossus had forewings that reached the length of 57 to 71 mm (2.2 to 2.8 in).[7] They possessed an inflated frons and a long rostrum (piercing and sucking mouthpart), indicating that they fed on xylem fluids like some other modern hemipterans.[8]
The host plants of palaeontinids have been assumed to be ginkgophytes based on the geographic distribution of both groups. The extinction of palaeontinids during Early Cretaceous has been linked to the decline of ginkgophytes at the end of the Mesophytic era (Late Permian to Middle Cretaceous) and the rise of angiosperms (flowering plants).[9][10] Numerous newly evolved insectivorous animals (feathered theropods, primitive mammals, and early birds) may have also contributed significantly to their extinction.[9]
Most species of palaeontinids exhibit cryptic coloration.[10] The patterns on their wings protected them as they perched on branches and fed on sap. They may also have served as secondary sexual characteristics. The color patterns can vary slightly within the same species.[8]
Palaeontinids, like modern cicadas, possess four membranous wings supported by veins. The length and width ratio of the wings can vary within the same species, sometimes as a result of fossil preservation.[8] Early Jurassic palaeontinids, like Suljuktocossus, exhibit the most primitive wing forms in the family.[11] The forewing was elliptical with the "nodal line" (the area where the wing bends during flight, also known as the "transverse flexion line") more or less dissecting through the center of the wing. The hindwing was short and broad. The bases of the forewings overlapped that of the hindwings like in modern butterflies. Taken together with their large bodies, these characteristics indicate that they were fast but moderately versatile fliers.[12]
In contrast, later palaeontinids like the Upper Jurassic Eocicada and Early Cretaceous Ilerdocossus had triangular forewings with the flexion line closer to the base. They had smaller and narrower hindwings that did not overlap with the forewing. These indicate that they were highly versatile fliers, able to fly with a wide range of speeds and agility like modern wasps and sphinx moths.[12] They also possessed changes to the leading edge of their forewings, suggesting an overall gain in lift.[11]
The trend of forewing elongation is most evident in members of the family Mesogereonidae, an early offshoot and close relatives of palaeontinids.[13]
Classification
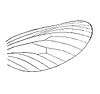
(forewing)
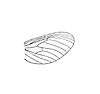
(forewing)
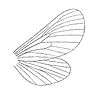
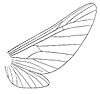
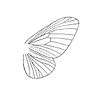
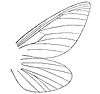
The family was first erected by the Austrian entomologist Anton Handlirsch in 1908. Like Butler, Handlirsch insisted that palaeontinids were members of lepidopteran Heteroneura (butterflies and moths). Palaeontinids were then only known mostly from poorly preserved specimens like Palaeontina and Eocicada. He claimed they were related to the extant family Limacodidae (slug moths).[14] The English entomologist Edward Meyrick supported the lepidopteran conclusion, though he believed they belonged to the family Hepialidae (ghost moths) instead. He said "There is little doubt that it [i.e. Palaeontina oolitica] belongs to the Hepialidae."[3]
The Belgian entomologist Auguste Lameere challenged this conclusion, claiming palaeontinids were more closely related to the extant family Cicadidae (cicadas). The English-Australian entomologist and geologist Robert John Tillyard supported Lameere's conclusion, noting that the wings of palaeontinid fossils lacked the characteristic scales of lepidopterans but instead had tubercules, pits, and cross-ridges like those found in modern cicadas.[14] He also cited characteristics of wing venation that distinctly differs from that of lepidopterans.[3]
Palaeontinidae are currently classified under the extinct superfamily Palaeontinoidea along with the families Dunstaniidae and Mesogereonidae.[11] They are classified under infraorder Cicadomorpha of the hemipterans (true bugs).[15]
The name Cicadomorphidae was once proposed as a replacement for the name Palaeontinidae in 1956 by the Australian entomologist J.W. Evans. This was because of Handlirsch's earlier insistence that the type species Palaeontina oolitica may not have been Hemipteran. However, Evans later conceded that retaining the name Palaeontinidae was preferable as the drawings Handlirsch based his conclusions on were from badly preserved specimens.[16]
Evolution
Riek (1976) originally considered Palaeontinoidea to be the descendants of the family Cicadoprosbolidae (currently known as the family Tettigarctidae), insects believed to be transitional between the ancestral cicada-like family Prosbolidae and the modern family Cicadidae.[11]
Wang et al (2009), however, notes that palaeontinoids more closely resemble prosbolids in agreement with earlier studies by Wootton (1971), Shcherbakov (1984), and Shcherbakov and Popov (2002). They conclude that palaeontinoids descended directly from the family Prosbolidae rather than from tettigarctids.[11] Modern cicadas therefore, did not descend directly from Palaeontinidae.
Within Palaeontinoidea, the family Dunstaniidae (Upper Permian to Lower Jurassic of Australia, South Africa, and China) is ancestral to palaeontinids. Both are distinct from the only other member of the superfamily, the more primitive and specialized family Mesogereonidae (Upper Triassic of Australia and South Africa).[11]
Distribution and geologic time range
.jpg)
Palaeontinids first appeared during the Rhaetian age of the Upper Triassic[17] and became extinct during the Early Aptian age of the Lower Cretaceous (203.6 to 112.0 million years ago).[9][15] They achieved their greatest diversity during the Jurassic period.[18]
The earliest palaeontinid discovered, is the poorly known genus Asiocossus, based on a forewing fragment recovered from the Upper Triassic of Kyrgyzstan.[17][19] Palaeontinid fossils are abundant in Eurasia and South America.[11] Fossils have been recorded in Brazil, China, Russia, Germany, the Transbaikal region, Tajikistan, Turkmenistan, Kyrgyzstan, Kazakhstan, Spain, and the United Kingdom. Important localities for palaeontinid fossils include the Crato Formation Lagerstätte of Brazil and the Yixian Formation, Haifanggou (or Jiulongshan) Formation, and the Daohugou Beds of China.[7][8][20]
Genera
The following is the list of genera classified under Palaeontinidae:[21]
- †AbrocossusWang & Zhang, 2007 in Wang et al. 2007a - Middle Jurassic, East Asia
- †ArchipsycheHandlirsch, 1906–1908 - Late Jurassic, Central Europe
- †AsiocossusBecker-Migdisova, 1962 - Upper Triassic, Central Asia[19]
- †BaeocossusMenon et al. 2005 - Early Cretaceous, Eastern South America
- †BeloptesisHandlirsch, 1906–1908 - Late Jurassic, Central Europe
- †CicadomorphaMartynov, 1926 - Late Jurassic, Central Asia; Early Cretaceous North Asia
- †ColossocossusMenon et al. 2005 - Early Cretaceous, Eastern South America
- †CratocossusMartins-Neto, 1998 - Early Cretaceous, Eastern South America
- †DaohugoucossusWang et al. 2006b - Middle Jurassic, East Asia
- †EocicadaOppenheim, 1888 - Late Jurassic, Central Europe
- †EoiocossusWang et al. 2006c in Wang et al. 2006c - (includes Papilioncossus Wang et al. 2007c) Middle Jurassic, East Asia
- †GansucossusWang et al. 2006b - Middle Jurassic, East Asia
- †HamicossusWang & Ren 2007ab - Middle Jurassic, East Asia
- †IlerdocossusGomez-Pallerola, 1984 - (includes WonnacottellaWhalley & Jarzembowski, 1985 and LiaocossusRen et al., 1998) Early Cretaceous, Western Europe & East Asia
- †LimacoditesHandlirsch, 1906–1908 - Late Jurassic, Central Europe
- †MartynovocossusWang & Zhang 2008 in Wang et al. 2008 - (= PseudocossusMartynov, 1931) Early to Middle Jurassic, North Asia; Late Jurassic, Central Asia
- †MiracossusRen et al. 1998 - Early Cretaceous, East Asia
- †MontsecocossusGomez-Pallerola, 1984 - Early Cretaceous, Western Europe
- †NeimenggucossusWang & Zhang, 2007 in Wang et al. 2007a - Middle Jurassic, East Asia
- †NingchengiaWang, Zhang & Szwedo, 2009 - (= Fletcheriana Evans, 1956 in partim) Middle Jurassic, East Asia
- †PachypsycheHandlirsch 1906 - Early Cretaceous, Western Europe
- †PalaeocossusOppenheim, 1885 - Early Jurassic, Central and North Asia
- †PalaeontinaButler, 1873 - Early Jurassic, Central and North Asia
- †PalaeontinodesMartynov, 1937 - (= IjacossusBecker-Migdisova, 1950; Shcherbakov 1985) Early to Middle Jurassic, Central and North Asia; Middle Jurassic, Central Asia
- †ParawonnacottellaUeda, 1997 - Early Cretaceous, Eastern South America
- †PhragmatoecicossusBecker-Migdisova, 1949 - Early to Middle Jurassic, Central Asia
- †PhragmatoecitesOppenheim, 1885 - Early to Middle Jurassic, North Asia
- †PlachutellaBecker-Migdisova, 1949 - Early to Late Jurassic, Central and Eastern Asia
- †ProlystraOppenheim, 1888 - Late Jurassic, Central Europe
- †ProtopsycheHandlirsch, 1906–1908 - Late Jurassic, Central Europe
- †ShurabocossusBecker-Migdisova, 1949 - Early to Middle Jurassic, Central Asia
- †SinopalaeocossusHong, 1983- (= Quadraticossus Wang & Ren, 2007a) Middle Jurassic, East Asia
- †SuljuktajaBecker-Migdisova, 1949 - Early to Middle Jurassic, Central Asia
- †SuljuktocossusBecker-Migdisova, 1949 - (= Fletcheriana Evans, 1956 in partim) Early Jurassic, Central Asia; Middle Jurassic, North Asia
- †TurgaiellaBecker-Migdisova & Wootton, 1965 - Jurassic, Central Asia
- †Valdicossus Wang, Zhang & Jarzembowski 2008 - Early Cretaceous, Western Europe
- †YanocossusRen, 1995 - Early Cretaceous, East Asia
- †CylloniumWestwood, 1854 - Early Cretaceous, Western Europe (too poorly preserved)[5][22]
- †PalaeontinopsisMartynov, 1937 - Early Jurassic, Central Asia (nomen dubium)
- †Palaeontinopsis sinensis- Hong, 1986; Zhang, 1997 Middle Jurassic, East Asia
- †Fletcheriana jurassica- Zhang, 1997
- †Fletcheriana magna- Riek, 1976
See also
References
- ↑ David M. Martill; Günter Bechly & Robert F. Loveridge (2007). The Crato fossil beds of Brazil: window into an ancient world. Cambridge University Press. pp. 284–287. ISBN 978-0-521-85867-0.
- ↑ "Family Palaeontinidae". The EDNA Fossil Insect Database. Retrieved July 16, 2011.
- 1 2 3 4 5 R.J. Tillyard (1919). "The Panorpoid Complex 3" (PDF). Proceedings of the Linnean Society of New South Wales (44): 533–718.
- ↑ Arthur Gardiner Butler (1869–1874). Lepidoptera Exotica; or, Descriptions and Illustrations of Exotic Lepidoptera (PDF). E. W. Jansen. pp. 126–127.
- 1 2 Bo Wang; Haichun Zhang & Edmund A. Jarzembowski (2008). "A new genus and species of Palaeontinidae (Insecta: Hemiptera: Cicadomorpha) from the Lower Cretaceous of southern England" (PDF). Zootaxa. Magnolia Press (1751): 65–68. ISSN 1175-5326. Retrieved July 13, 2011.
- ↑ Wang Ying & Ren Dong (2007). "Two new genera of fossil palaeontinids from the Middle Jurassic in Daohugou, Inner Mongolia, China (Hemiptera, Palaeontinidae)" (PDF). Zootaxa. Magnolia Press (1390): 41–49. ISSN 1175-5334. Retrieved July 15, 2011.
- 1 2 3 Federica Menon & Sam W. Heads (2005). "New species of Palaeontinidae (Insecta: Cicadomorpha) from the Lower Cretaceous Crato Formation of Brazil" (PDF). Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie). Staatliches Museum für Naturkunde (357): 1–10. ISSN 0341-0153. Retrieved July 15, 2011.
- 1 2 3 4 Bo Wang; Haichun Zhang; Yan Fang; Dejin Wang & Yutao Zhang (2007). "A revision of Palaeontinidae (Insecta: Hemiptera: Cicadomorpha) from the Jurassic of China with descriptions of new taxa and new combinations" (PDF). Geological Journal. Wiley InterScience. 43: 1–18. doi:10.1002/gj.1092. Retrieved July 15, 2011.
- 1 2 3 Bo Wang; Haichun Zhang; Yan Fang; Dejin Wang; Shengzhu Ji (2008). "New data on Cretaceous Palaeontinidae (Insecta: Hemiptera) from China" (PDF). Cretaceous Research. Elsevier (29): 551–560. doi:10.1016/j.cretres.2008.01.007. ISSN 0195-6671. Retrieved July 15, 2011.
- 1 2 D. E. Shcherbakov (2000). "Permian Faunas of Homoptera (Hemiptera) in Relation to Phytogeography and the Permo-Triassic Crisis" (PDF). Paleontological Journal. 34 (Suppl. 3): S251–S267. Retrieved July 15, 2011.
- 1 2 3 4 5 6 7 Bo Wang; Haichun Zhang & Jacek Szwedo (2009). "Jurassic Palaeontinidae from China and the Higher Systematics of Palaeontinoidea (Insecta: Hemiptera: Cicadomorpha)". Palaeontology. The Palaeontological Association. 52 (Part 1): 53–64. doi:10.1111/j.1475-4983.2008.00826.x.
- 1 2 Robin J. Wootton (2002). "Reconstructing insect flight performance from fossil evidence" (PDF). Acta zoologica cracoviensia. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences. 46 (suppl. – Fossil Insects): 89–99. ISSN 1734-915X. Retrieved July 15, 2011.
- ↑ R. Wootton (1971). "The Evolution of Cicadoidea (Homoptera)" (PDF). Proceedings: XIII International Congress of Entomology, Moscow (1): 318–319. Retrieved July 15, 2011.
- 1 2 R.J. Tillyard (1935). "The Evolution of the Scorpion-flies and their Derivatives (Order Mecoptera)" (PDF). Annals of the Entomological Society of America. Entomological Society of America. 28 (1): 1–45. Retrieved July 15, 2011.
- 1 2 "Palaeontinidae". Paleobiology Database. Retrieved July 15, 2011.
- ↑ J.W. Evans (1963). "The Phylogeny of the Homoptera" (PDF). Annual Review of Entomology. Annual Reviews. 8: 77–94. doi:10.1146/annurev.en.08.010163.000453. ISSN 0066-4170. Retrieved July 21, 2011.
- 1 2 Boris B. Rohdendorf; Donald Ray Davis, eds. (1991). Fundamentals of paleontology: Arthropoda, Tracheata, Chelicerata. Volume 9. Smithsonian Institution Libraries and the National Science Foundation. p. 220–224.
- ↑ Kyoichiro Ueda (1996). "A New Palaeontinid Species from the Lower Cretaceous of Brazil (Homoptera: Palaeontinidae)" (PDF). Bulletin of the Kitakyushu Museum of Natural History. Kitakyushu Museum and Institute of Natural History. 16: 99–104. Retrieved July 21, 2011.
- 1 2 Frank M. Carpenter & Roger L. Kaesler (1992). "Part R: Arthropoda 4". In Roger L. Kaesler & Raymond Cecil Moore. Treatise on Invertebrate Paleontology (PDF). Volume 3: Superclass Hexapoda. The Geological Society of America, Inc. & The University of Kansas. pp. 214–217. ISBN 978-0-8137-3019-6. Retrieved July 21, 2011.
- ↑ Xavier Martínez-Delclòs (1990). "Insectos del Cretácico inferior de Santa Maria de Meià (Lleida): Colección Lluís Maria Vidal i Carreras" (PDF). Treballs del Museu de Geología de Barcelona (in Spanish). Museo de Geología de Barcelona. 1: 91–116. Retrieved July 21, 2011.
- ↑ Mikko Haaramo (May 5, 2009). "†Palaeontinidae: After Grimaldi & Engel, 2005, Wang, Zhang & Fang, 2006, and Wang, Zhang & Szwedo, 2009". Mikko's Phylogeny Archive. Retrieved July 15, 2011.
- ↑ J.O. Westwood (1854). "Contributions to Fossil Entomology". The Quarterly Journal of the Geological Society of London. Geological Society of London. 10: 395–396. doi:10.1144/gsl.jgs.1854.010.01-02.43. Retrieved July 13, 2011.
|chapter=ignored (help)
External links
 Data related to Palaeontinidae at Wikispecies
Data related to Palaeontinidae at Wikispecies Media related to Palaeontinidae at Wikimedia Commons
Media related to Palaeontinidae at Wikimedia Commons